
MATSUBARA LAB

≫ PROFILE
| 2001年 | 大阪大学 歯学部歯学科 卒業 |
| 2005年 | 大阪大学 大学院歯学研究科 修了 |
| 大阪大学大学院歯学研究科 COE 博士研究員 | |
| 2008年 | Harvard school of Dental Medicine (Department of Infection and Immunity) Research fellow |
| 2012年 | 東北大学 矯正歯科 病院助教 |
| 2015年 | 九州歯科大学 健康増進学講座 分子情報生化学分野 助教 |
| 2019年 | 九州歯科大学 健康増進学講座 分子情報生化学分野 准教授 |
≫ LAB MEMBERS
KAZUMA YASUDA(安田 和真)…[Graduate Student, 大学院生]
KANA MIZUTA(水田 奏)…[Graduate Student, 大学院生]
HIROKA KAWAUE(川上 紘佳)…[Graduate Student, 大学院生]
≫ OUR RESEARCH
1.骨代謝研究
骨は物理的に体を支える組織で、歯科関連においても歯を支える歯槽骨の機能は重要です。また骨は筋肉の動きや様々な生体反応に欠かせないカルシウムの血中濃度を調節する機関です。血液中カルシウム濃度を調節するために、骨は常に破壊と形成(骨リモデリング)を繰り返しています。この骨の破壊は破骨細胞と呼ばれる細胞が、骨の形成には骨芽細胞と呼ばれる細胞が中心的な役割を担っています。破骨細胞と骨芽細胞はお互いにコミュニケーションをとりながら、血中カルシウム濃度と骨組織をうまく維持しています。このバランスが崩れると、骨粗しょう症や大理石骨病、口腔関連では歯周病などの病気になります。そのため、破骨細胞や骨芽細胞の機能を分子的に解明することを目的として研究を行っています。
1-1 破骨細胞
骨粗しょう症や歯周病など骨に関する代謝性骨疾患には、破骨細胞の機能が亢進する疾患が多く存在します。破骨細胞が骨吸収をおこなうには、「破骨細胞に分化する」、「破骨細胞が骨に接着する」、「破骨細胞が酸や基質分解酵素により骨基質を分解する」の3ステップを要します。これまで骨粗しょう症の治療薬として、破骨細胞のアポトーシスを誘導するビスホスフォネート製剤や破骨細胞の分化を抑制する抗RANKL抗体など、破骨細胞を無くして相対的に骨形成を優位にする薬剤がメインで使用されています。しかし、この治療法では歯科で問題視されている顎骨壊死などの有害事象が起きやすくなっています。これは、破骨細胞がないために骨リモデリングに異常をきたしているのが一因だと考えられています。そこで破骨細胞が存在したまま骨吸収機能を制御できるようになれば、有害事象が軽減された治療法の開発につながると考えられます。
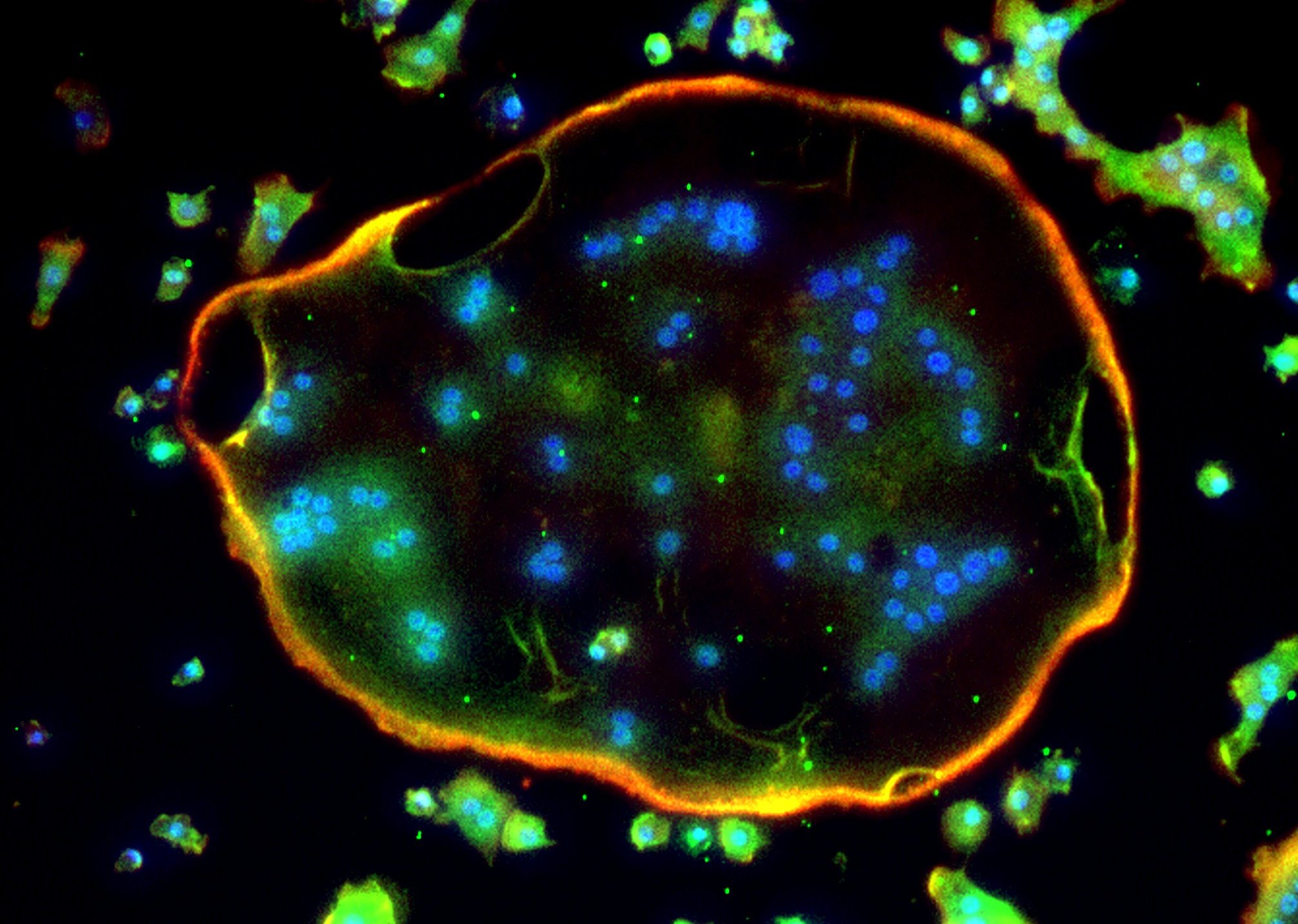
破骨細胞は骨基質に接着するためにアクチンリングという特殊な細胞骨格構造をとる。
私はこれまで破骨細胞の骨基質への接着に関して研究を中心に行ってきました。破骨細胞の細胞接着にはSrcというチロシンキナーゼが重要な働きをおこなっています。Srcノックアウトマウスは、破骨細胞が存在し、酸や基質分解酵素を産生する能力があるにもかかわらず、骨基質にうまく接着できずに骨吸収が行われないため、大理石骨病を呈します。このSrcがどのように破骨細胞の接着を制御しているのかを明らかにできれば代謝性骨疾患の治療法の発展につながるのではないかと考え、Srcを中心とした分子シグナルの解明を目的に研究を行っています。これまでにSrcの下流で働くCortactin (T. Matsubara, et. al., J. Bone Miner. Metab. 24, 368–372 (2006))、Srcの機能を調節するCbp/PAG (業績4)、PPP1r18 (業績2)、plectin (業績1, 3)に関する研究を発表しています。現在、さらに他の分子の関連を調べています。
1-2 骨芽細胞・軟骨細胞
骨の維持には破骨細胞だけではなく、骨芽細胞の機能も重要です。これまでの研究で、骨芽細胞分化に必須の転写因子Osterixの発現制御機構(業績5)の一端を解明しました。Osterixとは話が異なりますが、骨芽細胞は関連する細胞の一つである骨細胞と一緒にメカニカルストレスを感知する可能性が考えられています。特に歯科領域においては矯正治療時の歯の移動に大きく関わっています。メカニカルストレスは細胞骨格系のタンパク質に作用し、細胞内のシグナル分子を活性化しますが、そこにはまだ不明な点が多く残っています。現在、Src関連タンパク質と細胞内シグナルに関して研究を行っています。
また詳細は公表できませんが、骨・軟骨に症状の現れる難病の遺伝子についても研究を行っています。
2.がん研究
破骨細胞の細胞接着に重要なSrcは、がん原遺伝子として同定されたタンパク質で、がんで高いキナーゼ活性を有し、がんの成長、浸潤・転移に関わっているとされています。そこで破骨細胞研究で培った知識と技術を元に、Srcと関連タンパク質のがんにおける役割を検討しています。
≫ SELECTED PUBLICATIONS
[1] T. Matsubara, T. Yaginuma, W.N. Addison, Y. Fujita, K. Watanabe, I. Yoshioka, H. Hikiji, K. Maki, R. Baron, S. Kokabu, Plectin stabilizes microtubules during osteoclastic bone resorption by acting as a scaffold for Src and Pyk2, Bone. 132 (2020) 115209. https://doi.org/10.1016/j.bone.2019.115209.
[2] T. Matsubara, S. Kokabu, C. Nakatomi, M. Kinbara, T. Maeda, M. Yoshizawa, H. Yasuda, T. Takano-Yamamoto, R. Baron, E. Jimi, The Actin-Binding Protein PPP1r18 Regulates Maturation, Actin Organization, and Bone Resorption Activity of Osteoclasts, Mol. Cell. Biol. 38 (2018) e00425-17. https://doi.org/10.1128/MCB.00425-17.
[3] T. Matsubara, M. Kinbara, T. Maeda, M. Yoshizawa, S. Kokabu, T. Takano Yamamoto, Regulation of osteoclast differentiation and actin ring formation by the cytolinker protein plectin, Biochem. Biophys. Res. Commun. 489 (2017) 472–476. https://doi.org/10.1016/j.bbrc.2017.05.174.
[4] T. Matsubara, F. Ikeda, K. Hata, M. Nakanishi, M. Okada, H. Yasuda, R. Nishimura, T. Yoneda, Cbp recruitment of Csk into lipid rafts is critical to c-Src kinase activity and bone resorption in osteoclasts, J. Bone Miner. Res. 25 (2010) 1068–1076. https://doi.org/10.1359/jbmr.091039.
[5] T. Matsubara, K. Kida, A. Yamaguchi, K. Hata, F. Ichida, H. Meguro, H. Aburatani, R. Nishimura, T. Yoneda, BMP2 regulates Osterix through Msx2 and Runx2 during osteoblast differentiation, J. Biol. Chem. 283 (2008) 29119–29125. https://doi.org/10.1074/jbc.M801774200.